Jeśli przebrnąłeś, Drogi Czytelniku, przez moje poprzednie dwa artykuły (str. 3065 i 3150) i masz wrażenie jakbyś przedzierał się przez gęsto zarośniętą przez prebiotyczne stwory dżunglę — masz rację. Nauka o początkach życia to teren wciąż na nowo poznawany, niczym nietknięte ludzką stopą skrawki ziemi, na których botanik z zaskoczeniem odnajduje dziwnego rodzaju kwiaty. Trzymając się dalej tego porównania, wędrówka przez zagadnienie cząsteczek prebiotycznych i pierwszych systemów biochemicznych to łatwiejsza część naszej podróży. I nie chodzi tu o stopień skomplikowania czy też zaawansowania w posiadanej wiedzy - chcąc nie chcąc pchamy się w głębokie błoto z ruchomymi piaskami. Brak nam dobrego przewodnika, jest ich kilku, ale ich nabazgrane na owczej skórze mapy mówią zupełnie coś innego. Najgorsze jest to, że jak już wdepniemy w bagno, konsekwentnie musimy się przez nie przedrzeć, aby uniknąć wyśmiania przez prymitywne plemię Kreacjonusów (które od pokoleń jest zdania, że przez bagna przedzierać się nie warto). Zadanie to ciężkie, ale spróbuję nakreślić mały przewodnik dla odkrywców pierwszych zaczątków już jak najbardziej współczesnego nam życia.
Mimo,
iż
najbardziej podstawowa definicja życia twierdzi, że tworem żywym
jest układ
biochemiczny podlegający ewolucji darwinowskiej, jesteśmy raczej
skłonni
akceptować życie w formie komórkowej. To już jakby namacalny dowód -
możemy takie
„coś" obejrzeć pod mikroskopem i być świadkiem jak zawzięcie macha
rzęską w poszukiwaniu pokarmu. Żywa komórka nie jest jednak niczym innym niż
imponująco
skoordynowanym i skomplikowanym układem biochemicznym uwięzionym w błonie
komórkowej. Właściwie należałoby użyć słowa „chronionym", gdyż
błona
komórkowa nie jest bynajmniej nieprzenikalną ścianą, a bardzo złożonym,
selektywnie przepuszczającym tworem. Rycina 1 to bardzo duże
uproszczenie tego niesamowitego, dynamicznego tworu.
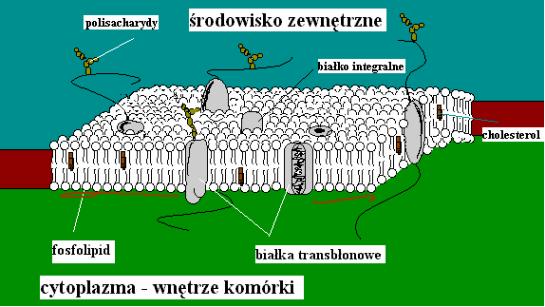
Rycina 1. Mozaikowy model błony współczesnych komórek.
Oprócz lipidów, składa się ona z wszelakiej maści białek transbłonowych biorących udział w transporcie cząsteczek do i z wnętrza komórki. Błona komórkowa jest przepuszczalna jedynie dla niektórych. Pośród nich znajdują się cząsteczki wody oraz gazów takich jak tlen, azot i dwutlenek węgla oraz dla lipidów takich jak cholesterol. Ograniczona przepuszczalność dotyczy niektórych jonów, cukrów i innych małych molekuł. Przepływ wody może być zwiększony poprzez ostatnio intensywnie badane kanały wodne. Przez miliardy lat błony wykształcały system kanałów do biernego transportu np. jonów, co jest podstawą wrażliwości komórek. Oprócz tego istnieje również wyrafinowany system transportu aktywnego (zużywającego energię chemiczną) — pozwalający na przenoszenie „pod prąd". Powrócimy do tego zagadnienia trochę później. Błona komórkowa wystawia swoje „czułki" w postaci łańcuchów cukrowych, co jest podstawą wszelakiej interakcji ze środowiskiem. Cała struktura jest elastyczna i znajduje się w ciągłym ruchu.
Jeśli chcemy być konsekwentni, musimy założyć kilka rzeczy, które pozwoliłyby na powstanie pierwszych tworów komórkopodobnych.
Po pierwsze: pierwotna błona musiała uformować się spontanicznie z materiałów dostępnych na młodej Ziemi, bądź/oraz zawleczonych z kosmosu. Mało prawdopodobne byłoby założenie, że pływające swobodnie cząsteczki utworzyły miejscowe zagęszczenie prowadząc do skomplikowanego systemu produkcji lipidów i ich dystrybucji wokół zagęszczenia.
Po drugie: błona ta oprócz oczywistej ochrony systemu biochemicznego od środowiska będzie zapewniała chociażby najbardziej podstawowy rodzaj transportu. Miałoby to służyć do utrzymania odpowiedniego stężenia jonów po obu stronach, dostarczania substratów do syntezy wewnątrz oraz usuwanie niepotrzebnych produktów.
Po trzecie: powstała błona musi umożliwić „rozmnażanie" pierwotnej komórce. Czy byłoby to prymitywne pączkowanie, czy bardziej lub mniej zaawansowany podział — sprawny układ biochemiczny musi przekazać korzystne cechy następnemu „pokoleniu".
Powstanie. Witamy w tłustym towarzystwie
Wydaje się całkiem prawdopodobne, że nie tylko cząsteczki dostępne na młodej Ziemi, ale również składniki pozaziemskich obiektów mogły być źródłem powstania pierwszych błon. Szacuje się, że bombardowanie meteorytami wczesnej Ziemi mogło dostarczyć około 107 kg na rok organicznych materiałów [1]. Wydaje się, iż rodzime surowce byłyby wystarczające, trudno jest jednak wykluczyć zawleczenia zarodków życia na Ziemię.
Wróćmy do budowy błony komórkowej, a właściwie do podstawowych „cegiełek" ją budujących. Ograniczę się jednak do podstawowych informacji, niezbędnych do dalszej lektury artykułu. Głównym składnikiem obecnych błon komórkowych są fosfolipidy (Rycina 4A) — cząsteczki zaliczane do grupy związków amfifilowych. Oznacza to, że posiadają obydwie cechy z reguły pojedynczo przypisane do jednej cząsteczki. W tym przypadku jest ona podzielona na dwa regiony — umowna "głowa" jest hydrofilowa, czyli „lubi" środowisko wodne, zaś "ogon" jest hydrofobowy, czyli będzie zawsze dążył od ukrycia się przed cząsteczkami wody.
Wyobraźmy sobie mieszaninę takich cząsteczek — otóż mają one skłonność do spontanicznego łączenia się w tzw. micele — drobne pęcherzyki oraz w dwuwarstwy (Rycina 2 i 3 — widać, że twory te powstają na skutek „uciekania" części hydrofobowych cząsteczek od środowiska wodnego - próbują się one upakować jak najciaśniej, wystawiając na zewnątrz części hydrofilowe). W specyficznych warunkach mogą również tworzyć zamknięte dwuwarstwy, na kilka różnych sposobów opisanych dalej.
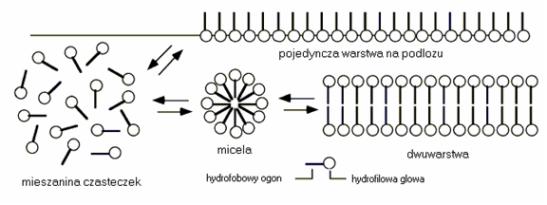
Rycina 2. Formowanie uporządkowanych struktur przez cząsteczki amfifilowe. [2]
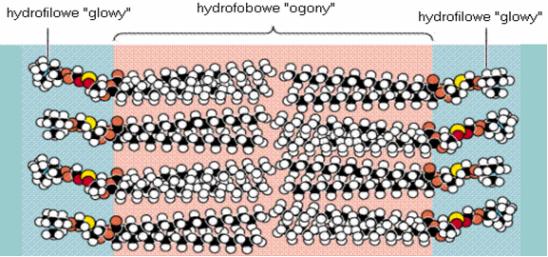
Rycina 3. Szczegółowy model dwuwarstwy utworzonej przez fosfolipidy.
Zadajmy więc podstawowe pytanie. Czy tak skomplikowane fosfolipidy były dostępne na prebiotycznej Ziemi? Najwyraźniej nie. Były jednak dostępne inne cząsteczki, znacznie prostsze lecz nadal zachowujące właściwości amfifilowe (spójrz na porównanie — Rycina 4). Wśród najpowszechniejszych znajdują się proste kwasy i alkohole oraz pochodne związków zwanych w skrócie PAH (polycyclic aromatic hydrocarbon — policykliczne węglowodory aromatyczne), proponuje się również pochodne izoprenów.
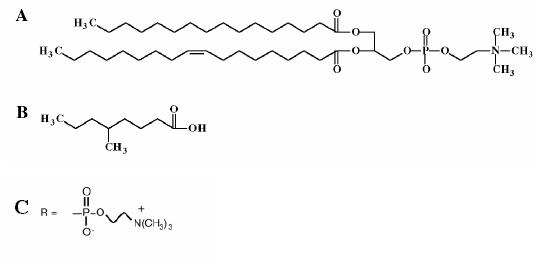
Rycina 4. Składnikiem współczesnych błon komórkowych są fosfolipidy (A). Składnikami pierwszych prymitywnych błon mogły być np. kwas 5-metylo nonanowy (B) obecny w meteorytach, czy substancja zwana w skrócie POPC (C). [2, 3]
O tym, że cząsteczki takiego rodzaju były zdolne do formowania zamkniętych dwuwarstw (pęcherzyków) świadczy doświadczenie zespołu dr Davida Deamera, który wyekstrahował je z skrawka meteorytu z Murchison. Ten dość spory kawałek gwiezdnej skały okazał się bezcennym znaleziskiem. Nie dosyć, że posiadał substancje uzyskane w eksperymencie Ureya-Millera (co potwierdziło jedynie nieuniknione powstanie życia we Wszechświecie), zawierał również cząsteczki amfifilowe takie jak kwas 5-metylononanowy (Rycina 4B) i przedstawicieli z grupy PAH. Otóż okazuje się, że takie cząsteczki mogą formować twory komórkopodobne w miejscowych zagęszczeniach [2]. Tutaj przychodzi z pomocą teoria lagun Stanleya Millera, o której wspominałem w poprzednich artykułach. Otóż parująca woda z niecek, brzegów mogła powodować zwiększenie stężenia w tym również omawianych przez nas cząsteczek. Tak więc podczas symulacji odpowiednio zagęszczone substancje amfifilowe spontanicznie formowały pęcherzyki (Rycina 5 A). Kalifornijscy naukowcy wskazują właśnie na PAH jako główne ich składniki. Mało tego — podobne doświadczenie przeprowadził dr James Dworkin używając jednak ekstraktu z zamrożonej materii międzygwiezdnej [2]. Tutaj głównym składnikiem była mieszanina kwasu i alkoholu dekanowego. (Rycina 5 B)
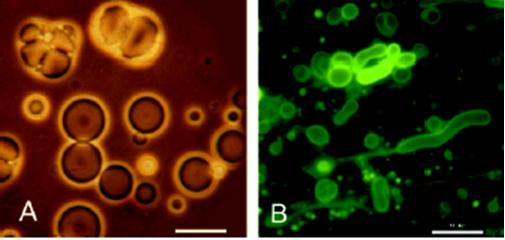
Rycina 5. (A) — ekstrakt z meteorytu z Murchison; pęcherzyki są zdolne do uwięzienia fluorescencyjnego barwnika. (B) — ekstrakt z materii międzygwiezdnej. Fluorescencyjna substancja barwi dwuwarstwy. [2, 3]
Spontaniczne formowanie się pęcherzyków nie wymaga zewnętrznej chemicznej energii, jednak zachowanych musi być kilka warunków. Jednym z nich jest, jak widzimy powyżej, ich odpowiednie zagęszczenie.
Proces ten mógł zachodzić na różne sposoby, takie jak wspomniane wyparowywanie w lagunach, krótkotrwałe zamarzanie i topnienie, czy adsorpcję na powierzchniach mineralnych [2]. Ostatni przypadek byłby najbardziej faworyzowany, gdyż, jak pamiętamy, na takich powierzchniach mogą być formowane katalitycznie aktywne RNA. Wnikanie cieczy z substancjami w szczeliny dawałoby bardzo dobrą ochronę przed nieprzyjaznymi warunkami zewnętrznymi. Okazuje się, że dwuwarstwowe pęcherzyki mogą być łatwo tworzone z miceli w cyklach ogrzewania i chłodzenia [4], które mogły być bardzo powszechnym zjawiskiem geologiczno-klimatycznym na młodej Ziemi. Proponuje się również inną możliwość — polimeryzację atmosferycznego metanu pod wpływem promieniowania UV. Substancja ta tworzyłaby swoistego rodzaju „kożuch" hydrofobowy pływający na dużych powierzchniach zbiornika i tworzyła parasol ochronny dla pływających pod nim cząsteczek [5]. Ostatnio ta kwestia stała się bardzo ważna, gdyż ponownie odkryto ślady komórkowych organizmów fotosyntetycznych datowanych na 3,7 miliarda lat, więc znacznie starsze zalążki życia musiały powstawać w wysoce niekorzystnych warunkach [6].
Przełomowe badania M. Hanczyca, S. Fujikawy i J. Szostaka publikowane w zeszłym roku zademonstrowały ewidentnie spontaniczne formowanie się dwuwarstwowych pęcherzyków w obecności ilastego minerału — montmorylonitu [7]. Patrz Rycina 5.1.
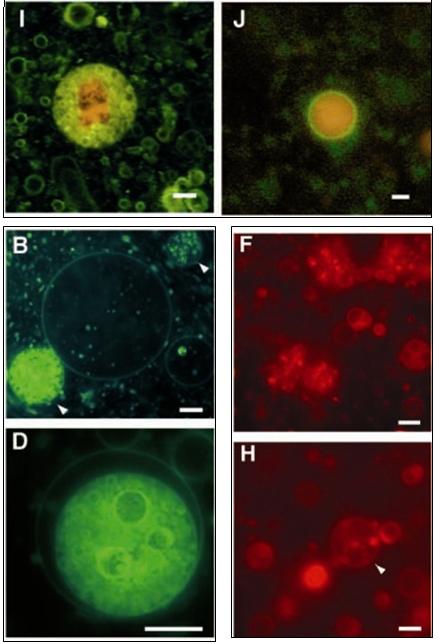
Rycina 5.1. Oto seria doświadczeń przeprowadzonych przez Hanczyca, Fujikawę i Szostaka. [7] Przedstawione tu pęcherzyki zostały uwidocznione fluorescencyjnie. Panele I, J przedstawiają czerwono barwione RNA zamknięte w wybarwionych na zielono pęcherzykach. Panele F,H przedstawiają wybarwiony montmorylonit — minerał wspomagający formowanie się pęcherzyków. Panele B,D pokazują pęcherzyki zawierające ciekawe twory aluminowo-krzemowe. [ 1 ]
Złapanie życia w ryzy
Powyżej wspomniana publikacja, moim zdaniem, jest równie wartościowa i sensacyjna jak pierwsze eksperymenty Stanleya-Millera (opisywany w pierwszym artykule). Naukowcy elegancko udowodnili, że formujące się pęcherzyki mogą zamykać w swoim wnętrzu gotowe RNA oraz fragmenty montmorylonitu, na których wciąż tworzą się nowe cząsteczki kwasu rybonukleinowego. Twory takie na dodatek mają zdolność do biernego, podstawowego transportu [7] — zaobserwowano z czasem przenikanie barwnika do wnętrz tych swoistych „pre-komórek".
P. Monnard, współpracujący z D.Deamerem, wykazał, że pęcherzyki formowane z kwasu i alkoholu dodekanowego mogą uwięzić mieszaninę cząsteczek DNA o długości około 600 par zasad (par nukleotydów) [2, 3]. Spójrz na poniższy rycina.
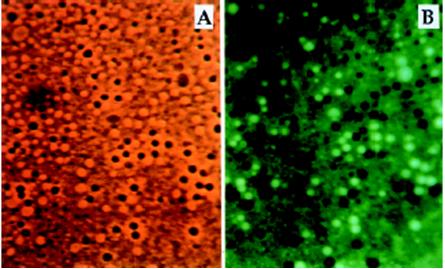
Rycina 6. Uwięzione DNA w pęcherzykach zbudowanych z kwasu i alkoholu dodekanowego. Pęcherzyki z DNA w panelu (A) są czarne, pod wpływem światła UV emitują intensywnie zielone światło. [3]
RNA uwięzione w takim pęcherzyku miało większą szansę na stworzenie funkcjonalnego systemu biochemicznego, niż pływając swobodnie w roztworze. Zwiększa się w tym wypadku możliwość wspólnych interakcji i nabycia nowych funkcji. Zwiększa się stopień uporządkowania całego układu. Co więcej, aktywowane nukleotydy mogły dyfundować w obydwie strony poprzez błonę, dostarczając budulca do nowych łańcuchów RNA. Inną drogą nukleotydów do wnętrza pre-komórki były naturalnie występujące, krótkotrwale tworzące się przerwy w dwuwarstwie. Zagadnienie transportu stanowi poważny problem, ograniczając możliwości pierwszych systemów metabolicznych.
Zakładamy raczej, że energia chemiczna była głównym źródłem zasilania w świecie prebiotycznym. A co z fotosyntezą? Czy ostatnie doniesienia o jej śladach 3,7 miliarda lat temu nie dają do myślenia? Aby wykorzystać energię słoneczną, fotony muszą być najpierw zaabsorbowane przez substancje pigmentowe i przekształcone w niestabilne formy energii chemicznej. Oczywiście systemy porównywalne do dzisiejszych komórek były jeszcze nieobecne, ale istniały wspomniane wcześniej związki z grupy PAH, które mogły zostać inkorporowane w zamknięte struktury dwuwarstwowe. Dotychczas pojawiło się kilka sygnałów o właściwościach PAH jako pigmentu [3]. Badania trwają.
Dzisiejszy, wydajny, nawet bierny (zgodnie z gradientem stężenia przenoszonej substancji) transport oparty jest na białkach przenośnikowych. (Rycina 7). Świat komórkowy dysponuje również fascynującym i wyrafinowanym systemem transportu aktywnego (wbrew gradientowi stężęń — Rycina 8) na wiele sposobów regulowanym.
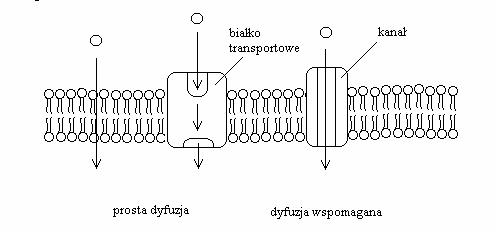
Rycina 7. Rodzaje transportu biernego współczesnych błon. Transport zachodzi zawsze zgodnie z gradientem stężeń przenoszonej cząsteczki - jeśli substancji jest więcej na zewnątrz, będzie ona transportowana do wnętrza, aby wyrównać stężenia.
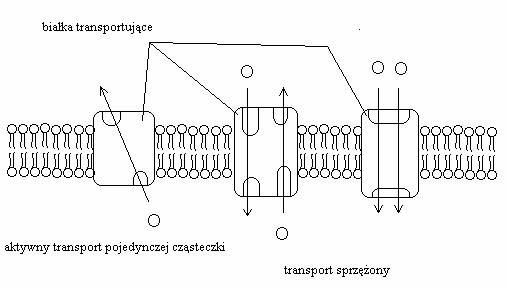
Rycina 8. Podstawowe rodzaje transportu aktywnego współczesnych błon. Transport zachodzi niezależnie od gradientu stężeń przenoszonych cząsteczek, ale wymaga dostarczenia energii. Innymi formami transportu aktywnego są również tutaj nie opisywane endo- i egzocytoza.
Możemy spodziewać się więc, że wysoka efektywność przenoszenia rozpoczęła się dopiero po pojawieniu się pierwszych białek, które mogły być „złapane" w błonie. Nie zaobserwowano RNA tworzącego twory transbłonowe. Jak to się stało — jest dla nas na razie nierozwiązaną zagadką, ale trzymajmy kciuki za biologów symulujących powstanie życia...
Czas na dzieci. Czas na lepszą ewolucję
Wreszcie dotarliśmy do kwestii przekazania materiału genetycznego. Wrócimy tutaj do wcześniej wspominanego artykułu trójki naukowców. Postawili oni następującą tezę: prymitywne rozmnażanie najprawdopodobniej polegało na prostym podziale pęcherzyka razem z materiałem zawartym w środku. Jeśli wewnątrz pęcherzyka wykształci się pewien jednolity system, podzielenie go na pół da nam dwa pęcherzyki z systemami zdolnymi do niezależnych zmian i niezależnej ewolucji. Jednak aby utrzymać ten podział takie pra-komórki powinny rosnąć. Okazało się, że pęcherzyki rosną pod wpływem zwiększania się ilości miceli w roztworze. Czyli najpierw powstają micele, które przekształcają się w nowe dwuwarstwowe pęcherzyki, po czym po pewnym wysyceniu ogranicza się powstawanie nowych pęcherzyków a zwiększająca się wciąż ilość miceli. Cześć składników miceli — w na razie szczegółowo nie poznany sposób — łączy się z pęcherzykami zwiększając ich objętość. Proces ten dotyczy 90% puli pęcherzyków w roztworze [7].
Jak zachodzi podział? Otóż bynajmniej nie poprzez reakcje biochemiczne. Naukowcy przepuścili podrośnięte pęcherzyki przez porowaty materiał (o porach rozmiarów 100 nm) i okazało się, że podzieliły się ona na pół z bardzo małą stratą zawartego materiału. Następnie, aby potwierdzić produktywność tego systemu, przeplatali cykle wzrostu (dodając micele) oraz podziału (przepychając przez wspomnianą „gąbkę"). Całe doświadczenie dowiodło, że wzrost pęcherzyków jest możliwy dzięki naturalnemu wzrostowi stężenia miceli, zaś podział może zaistnieć dzięki prostym procesom fizykochemicznym [7].
P. Monnard i D. Deamer poszli o krok dalej — zaproponowali minimalny model komórki oparty na katalitycznym i strukturalnym RNA zobrazowany na Rysunku 9.
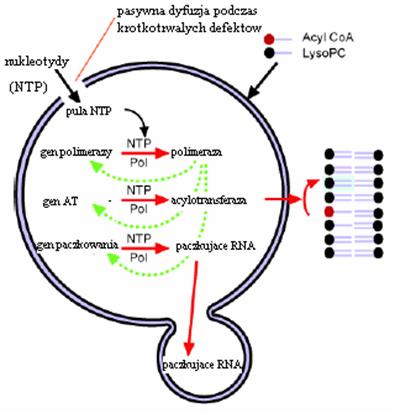
Rycina 9. Minimalny model komórki oparty na katalitycznym i strukturalnym RNA. Opis w tekście. [9]
Mimo iż teoria fragmentów RNA o właściwościach polimerazy RNA (produkowania kopii RNA; zob. poprzednie artykuły) jest niekompletna, to jest całkiem prawdopodobna i rozwijana [8]. Naukowcy twierdzą, że niezbędne byłyby trzy systemy [3]. Jeden zawierałby gen polimerazy, na podstawie którego polimeraza produkowałaby swoje bliźniacze kopie. Enzym ten produkowałby również na podstawie innego genu acylotransferazę, która aktywnie brałaby udział we wzroście błony komórkowej. Wreszcie namnażałby cząsteczki RNA odpowiedzialne za "pączkowanie". Model nie ma na razie poparcia doświadczalnego.
Jakimkolwiek torem podążał rozwój komórek — dzisiejsze efekty są imponujące. I pomyśleć, że około czterech, może i więcej miliardów lat temu wystarczyła iskierka. Resztę „roboty" wykonała ewolucja.
Ufam, że pomogłem Czytelnikom Racjonalisty nieco przybliżyć obecny stan wiedzy o początkach życia na Ziemi. Tutaj na razie kończymy naszą wędrówkę, mimo że na horyzoncie widać wiele terenów do eksploracji. Może kiedyś zaproszę jeszcze na podobną eskapadę w nieporównywalnie bardziej skomplikowany i równie fascynujący świat współczesnych komórek i pra-organizmów.
Dzisiejsza nauka potrafi przyprawić nas o zachwyt i coraz częściej zawroty głowy. Cieszę się ogromnie, że jestem świadkiem tak rewolucyjnych odkryć jak zeszłoroczny sukces Hanczyca, Fujikawy i Szostaka. Kilka tygodni temu koreańscy naukowcy, adaptując komórki macierzyste do hodowli, dokonali pierwszego zdecydowanego kroku w kierunku inżynierii tkankowej. Fizycy są coraz bliżej unifikacji, zwieńczenia ludzkiej nauki. Czekamy w napięciu na sygnały życia na Marsie. Mam nadzieję, że udało mi się chociaż trochę zarazić Czytelników moim entuzjazmem i zachęcić ich do własnych poszukiwań i rozmyślań nad początkami życia.
Literatura:
[1] Love, S.G. and Brownlee, D.E
(1993) „A direct measurement of the terrestrial mass accretion rate of
cosmic
dust". Science 262, 550-553.
[2] D. Deamer, J. P. Dworkin,
S.A.Sandford, M.P. Bernstein, and L.J. Allamandola, „The First Cell
Membranes",
Astrobiology, Volume 2, Number 4, 2002.
[3] P.A. Monnard, D.W.Deamer,
„Membrane Self-Assembly Processes: Steps Toward the First Cellular
Life", The
Anatomical Record, 268:196-207 (2002).
[4] Goltsov AN, Barsukow LI.,
„Synergetics of the membrane self-assembly: a micelle-to-besicle
transition", J
Biol Phys 2002; 26:27-41.
[5] Morchio R, Traverso s., „The
hydrophobic superficial layer: the primordial cradle of life?", Biology
Forum 1999, 92:105-117.
[6] Rosing MT, "13C-Depleted carbon
microparticles in >3700-Ma sea-floor sedimentary rocks from west
[7] M.M Hanczyc, S.M. Fujikawa, J.W.
Szostak, „Experimental Models of Primitive Cellular Compartments:
Encapsulation, Growth, and Division", Science, 2003, Vol
302:618-622.
[8]
Rysunki 2, 4, 5, 6 i 9 przerysowane, przetłumaczone i zamieszczone w artykule za zgodą dra Davida Deamera.
UWAGA: Czytelnicy mogą przeglądać oraz pobierać powyższy materiał tylko w celu prywatnego, niekomercjalnego użytku. Powyższy materiał jest chroniony przez prawo, nie może być reprodukowany, rozpowszechniany, modyfikowany, adaptowany, przedstawiany albo sprzedawany w całośći bądź w częściach bez uprzedniego zezwolenia wydawcy.
Marcin Klapczyński Ukończył biologię molekularną na Uniwersytecie Adama Mickiewicza w Poznaniu. Pracował jako Research Specialist in Health Science w Department of Anatomy and Cell Biology na University of Illinois w Chicago. Obecnie pracuje jako Associate Cell Biologist / Histologist w Abbott Laboratories (Illinois). Specjalizuje się w ekspresji białek 'od zera', hodowlach linii komórkowych, symulacji in vitro procesów zachodzących w komórkach. Jego pasją jest teoria ewolucji, w szczególności ewolucja systemów biochemicznych i pochodzenie życia we Wszechświecie. Ukończył biologię molekularną na Uniwersytecie Adama Mickiewicza w Poznaniu. Pracował jako Research Specialist in Health Science w Department of Anatomy and Cell Biology na University of Illinois w Chicago. Obecnie pracuje jako Associate Cell Biologist / Histologist w Abbott Laboratories (Illinois). Specjalizuje się w ekspresji białek 'od zera', hodowlach linii komórkowych, symulacji in vitro procesów zachodzących w komórkach. Jego pasją jest teoria ewolucji, w szczególności ewolucja systemów biochemicznych i pochodzenie życia we Wszechświecie.Liczba tekstów na portalu: 22 Pokaż inne teksty autora Liczba tłumaczeń: 1 Pokaż tłumaczenia autora |
Oryginał.. (http://www.racjonalista.pl/kk.php/s,3272)
(Ostatnia zmiana: 07-04-2004)